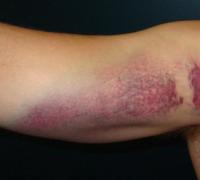
Как выглядит гренландский тюлень. Детеныши гренландских тюленей не могут выжить в условиях глобального потепления
Как бы ни были разнообразны экосистемы по величине, насыщенности видами, скорости создания и расходования продукции и в целом интенсивности круговорота, все они без исключения подчиняются определенному количественному соотношению первичной и вторичной продукции, называемому правилом пирамиды продукции. Это значит, что за единицу" времени на предыдущем трофическом уровне производится большее количество биомассы, чем на последующем. Если количество продукции, создаваемое на каждом трофическом у"ровне, изобразить в виде прямоугольников равной высоты и поставить их друг на друга, то быстро уменьшающаяся длина прямоугольников как раз и позволит представить эту" конструкцию в виде резко суживающейся кверху пирамиды. Пирамида продукции отражает закон расходования энергии в пищевых цепях.
В большинстве наземных экосистем действует также правило пирамиды биомасс, означающее, что суммарная фитомасса больше, чем биомасса фитофагов, или растительноядных организмов. Масса же растительноядных, в свою очередь, превышает биомассу хищников. Годовой прирост растительности по отношению к ее биомассе в наземных экосистемах невелик. Наиболее низки темпы прироста биомассы в лесах, где годовая продукция достигает лишь 2-6% от общей массы растений, сосредоточенной в крупных ветвях и стволах долгоживущих деревьев. Лишь в травянистых сообществах годовая продукция может достигать 40-55% биомассы (рис. 5.15). />Как следует из этих примеров, скорость создания органического вещества не определяетего суммарные запасы. Наличная биомасса организмов на каждом трофическом уровне в конкретных экосистемах зависит от того, как соотносятся между собой темпы накопле-
Рис. 5.15. Пирамиды биомасс некоторых биоценозов (по: Дрс, 1976).
Пр - продуценты, РК - растительноядные консумен- ты, ПК - плотоядные консументы, 3 - зоопланктон. Ф - фитопланктон. Цифры - биомасса сухого вещества в г/м2
лого медведя). Общий запас сайки в 1969 г. оценивался в 1,1 млн т. Ежегодное (1970-1973 гг.) изъятие одной трети биомассы быстро привело к сокращению его в 100 раз.
Огромное значение для всей морской баренцевоморской фауны имеет мойва. Во время подхода мойвы к берегам на нерест ее сопровождают стаи трески. В целом весь характер промысла трески определяется численностью, распределением и поведением мойвы. Икра мойвы имеет большое значение в питании пикши. В августе- октябре на севере Баренцева моря мойвой питаются тюлень, полярные акулы и крупная треска. Во время нерестовых миграций косяки мойвы сопровождаются также стаями морских птиц, в жизни которых она имеет большое значение.
Именно с баренцевоморской мойвой, как писал Г. Г. Матишов (1992), был допущен особенно крупный рыбохозяйственный просчет в 80-е гг. На судьбе популяции этой рыбы наиболее ярко отразилась хищническая практика современного промысла. Мало того, что интенсивно вылавливалась неполовозрелая популяция, была введена еще и машинная сортировка уловов, в результате которой мойва длиной менее 11 см отбраковывалась и, естественно, промысловой статистикой не учитывалась (статистикой учитывается только продукт - то. что заморожено траулером)- Об этом стоит сказать специально.
чтобы еще раз показать вопиющую бесхозяйственность в использовании ресурсов. Матишов приводит рассказ одного из штурманов мурманского тралового флота. Сельдь и мойва во время нереста собираются в огромные косяки. В таких районах всего при часовом тралении улавливается до 40 т. а иногда до 500-800 т. Большой морозильный рыболовный траулер в сутки может заморозить 30-40 т свежей рыбы. Избыток рыбы перерабатывается в муку или сбрасывается за борт. Сброшенная за борт рыба, как показывает практика, составляет не менее трети улова.
Быстрому падению запасов мойвы в конце 70-х-начале 80-х гг. способствовал не только пресс индустриального промысла, но и естественный процесс выедания ее треской и другими хищниками. Вследствие перелова сельди и сайки мойва стала основным пищевым ресурсом для гренландского тюленя, численность только беломорской популяции которого в те годы достигала 1 млн голов. Годовое потребление мойвы треской составляло 2-3 млн т.
В 70-е-80-е гг. баренцевоморское стадо трески находилось в депрессивном состоянии, и экосисистема откликнулась на сокращение пресса главного хищника вспышкой численности мойвы. Рыбаки немедленно усилили промысел, вылавливая примерно 4 млн т в год, т. е. две трети биомассы этой рыбы. Вследствие этого уже в 1986 г. угроза полного исчезновения мойвы как биологического вида стала очевидной и явной. Несмотря на введение полного запрета промысла пелагических рыб мойвы и сельди, численность их не восстанавливается до сего дня. Интенсивное изъятие мойвы резко ускорило и деградацию популяций баренцевоморекой трески.
Деградация птичьих базаров. Резким нарушением трофических связей в Баренцевом, а также в Белом морях вызваны и беды орнитофауны. Морские колониальные птицы, массовое скопление которых называют птичьими базарами, являются важной составной частью экосистемы Баренцева моря. Существование большого количества птиц обеспечивалось доступностью богатых рыбных кормов (мойва, сельдь, сайка, песчанка), обилием планктона и бентосных беспозвоночных в прибрежных водах и на литоралях.
Благодаря колониальному гнездованию значительную промысловую ценность представляли гага и кайра. В 30-60-е гт. их колонии на Новой Земле были главным районом охоты. Кайры достигают половой зрелости втрехлетнем возрасте. Сбор их крупных яиц массой в 100-120 г, обладающих прочной скорлупой и высокой питательностью, привел в конечном итоге к резкому сокращению кайровых базаров. Кроме -этого производился недопустимый отстрел и отлов кайр для заготовки корма ездовым собакам. В современный период численность птиц резко сократилась не только из-за истребления, но и из-за изъятия основной пиши - мелких рыб, а также неблагополучной зимовки 1986-1987 гг., когда в Баренцевом море погибло до 90% тонкоклювых и половина толстоюповых кайр. С 1988 г. наблюдается крайне низкий уровень размножения этих птиц на фоне массовой смертности взрослых особей.
На примере больших и серебристых чаек, наблюдения за динамикой численности которых проводились на Айновых островах, по казано, что в течение 60-80-х гг. происходил постоянный спад плотности популяций этих птиц. Почти во все сезоны отмечалась стопроцентная смертность птенцов от истощения. В 1981 и 1987 гг. корма уже не хватало взрослым птицам, и они тоже гибли. От 40 тыс. колониальных птиц здесь осталось около 2 тыс, особей. Началось формирование негнездовых скоплений в городской зоне. Тысячи голодных чаек появились в городах и поселках Мурманской и Архангельской областей.
Деградация птичьих базаров - это не только разрушение одного из верхних уровней трофической цепи. Это и снижение биопродуктивности морских экосистем, пока количественно не оцененное. Еше в середине 60-х гг. сотрудник Мурманского морского биологического института А. Н. Головкин показал существование тесной обратной связи между" морскими птица\ш и гидробионтами через обогащение прибрежных вод органическим веществом. П о его расчетам, в прибрежной зоне Баренцева моря птицы съедали в весенний (гнездовой) период примерно 70-80 тыс. т рыбы. Можно представить, сколько органики и фосфора они возвращали морю, удобряя его и обеспечивая развитие первопродуцентов. Резкое сокращение численности массовых видов пернатых, несомненно, приведет к разрыву обратной связи между гидробионтами и морскими птицами. Вне всякого сомнения, изменятся и скорость круговорота веществ и биопродуктивность экосистем. Точных расчетов пока нет. но есть все основания для тревога, ибо даже при прекращении промысла мойвы, сайки и сельди восстановление запасов этих стайных пелагических рыб проблематично.
Распад популяций морских млекопитающих. В высших звеньях экосистемы Арктики особую роль играют морские млекопитающие, самыми типичными из которых являются гренландский тюлень, морж, нерпа, морской заяц, хохлач. У берегов Кольского полуострова и арктических островов встречаются стада белух, в Баренцевом море мигрируют малый полосатик и другие беззубые киты и самые хищные из дельфинов - косатки. Гренландский тюлень в пищу использует пелагических беспозвоночных, мойву, сайку; белуха пита
ется преимущественно сайкой; моржи в основном поедают донных моллюсков.
Динамика численности этих морских млекопитающих за последние полвека определялась главным образом специализированным отечественным и норвежским промыслом. Многолетняя добыча поставила некоторые виды на грань почти полного исчезновения. За 1938-1964 гг., например, норвежцы выловили 78 тыс. мелких китов (в основном малых полосатиков - 93%). Регулярный интенсивный промысел в районах устойчивых скоплений китов закончился резким спадом их численности и свертыванием добычи.
Столь же трагична и судьба гренландского тюленя. Лишившись сайки - основного корма в период миграции, - гренландские тюлени беломорской популяции были вынуждены сменить схему миграции и с середины 70-х гг. стали появляться у северных берегов Кольскою полуострова в период нереста мойвы. Однако в середине 80-х гг. резкое ухудшение кормовой базы привело к распаду" и этих кормовых миграций. Тюлени стали оставаться вблизи берегов Мурмана на все лето. В 1986-1987 гг. с начала зимы около полумиллиона гренландских тюленей ушли к фьордам Норвегии, где примерно 60 тыс. погибло от истощения. \
Ухудшились и другие экологические условия существования тюленей. У некоторых из них. например, в желудке находится такое количество проглоченных пластиковых предметов, что звери не могут нормально переваривать пищу и обречены на табель.
Дисбаланс в биологических процессах экосистемы моря. Деградация верхних трофических уровней экосистемы привела к невостребованное™ его нижних этажей. В 60-80-е гг. ежегодное потребление эвфаузиид треской составляло 4 млн т, пикшей - 1 млн т и мойвой - 30 млн т. С учетом межгодовой изменчивости запасов этих рыб ежегодное потребление рачков колебалось в пределах 20-45 млн т (Дро- бышева, 1988. Цит. по: Матишов, 1992). Резкое сокращение во шорой половине 80-х гг. запаса этих промысловых рыб явилось одной из главных причин повышения (примерно и 4 раза) численности эвфаузиид. Однако сложившаяся пищевая цепь зоопланктон-хищные рыбы намного короче, чем трансформация энергии через пелагических рыб, что приводит к недоиспользованию продукции низких трофических уровней и уменьшению морской продуктивности. Важно отметить, что зоопланктон (эвфаузииды и калянус) может обеспечивать полноценное питание трески только до размера 20-30 см. Более крупным особям требуется и более энергоемкая и крупная пиша.
Эвфаузииды, а также калянус, являвшиеся кормом пелагических рыб, в настоящее время не утилизируются. Величина этой неиспользованной пищи, по-видимому, может достигать 50-60 млн т. Таким образом, при сложившейся в настоящее время ситуации огромное количество продукции низких трофических уровней не используется, что приводит к уменьшению биологической продуктивности водоема в целом. Следовательно, в Баренцевом море нарастает так называемое биологическое загрязнение.
В связи с изменением соотношений между планктоном и пелагическими рыбами в экосистемах возникла еще одна малоизученная ситуация: практическое исчезновение обратной связи, существовавшей в цепи продуктивность планктона-рыбопродуктивность, суть которой заключается во влиянии нереста, смертности и друтих проявлении жизнедеятельности рыб на развитие планктона. Например, еще недавно стада мойвы, с биомассой половозрелой части в 3- 4 млн т, поставляли в воду" около 55,2 тыс. т органического вещества. Резкая деградация птичьих базаров также снизила поступление питательных веществ для фитопланктона. Роль описанных масштабных явлений еще не оценена и только поставлена на повестку"дня.
Нет оснований ждать, что истощенные до предела популяции сельди, мойвы, сайки и трески смогут сами восстановиться. Кроме того, динамика экосистем начинает приобретать такие черты, при которых в Баренцевом море станут до минировать новые виды ихтиофауны, а истощенные промыслом рыбы-аборигены уже не смогут соперничать с пришельцами. Известно, например, что конкурентом сельди в питании в летнее время в юго-западной части моря является гребневик Bolinopsis, он скапливается здесь иногда в громадных количествах, превышающих 200 мг/м3. В таких случаях гребневики могут выедать калянуса, оттесняя сельдь в другие места. Резкий подъем численности гребневиков в 80-е гг. является, по-видимому, существенным препятствием восстановлению численности атлантическо- скандинавской сельди. В Норвежском море экологическую нишу"северного стада сельди прочно заняли путассу и отчасти анчоу"сы.
Таким образом, в Баренцевом море с момента подрыва запасов мойвы экосистема стала работать неэффективно с точки зрения как биологических законов, так и хозяйственных интересов.
Исследования биологических ресурсов дальневосточных морей. Необходимость экосистемного подхода в рыбохозяйственных исследованиях была осознана прежде всего при изучении пресных водоемов, где в первую очередь проявилось влияние хозяйственной деятельности человека. В 70-х гг. стало ясно, что и управление морскими биологическими ресурсами немыслимо без знания ключевых связей. определяющих функционирование экосистем. В начале 80-х гг. лаборатория прикладной биоценологии ТИНРО начала исследования морских экосистем во всей Д;ыьневосточной экономической зоне
Состав (%) и биомасса донных рыб в некоторых шельфовых районах
дальневосточных морей в 1980-е гг.
(по: Шунтов, 1995)
| Группа рыб | Район шельфа |
|||
| Западно- камчатский | Карагипско- олюторский | Северо- охотоморский | Берингов пролив |
|
| Сотовые | 0,9 | 0,2 | 10,1 | |
| Тресковые | 31,7 | 81,8 | 19,9 | 17,7 |
| Вельдюгогые | 0,3 | 1,0 | 6,2 | 3,2 |
| Рогатковыс (бычки) | 14,5 | 17,6 | 6,4 | 67,7 |
| Лисичковме | 0,9 | 0,3 | 0,1 | - |
| Ли паровые | 0,5 | 0,1 | 9,5 | 4,8 |
| Камбаловые | 51,0 | 8,7 | 37,7 | 5,0 |
| Прочие | 0,2 | 0,3 | 10,1 | 1,6 |
| Всего, тыс. т | 1075 | 615 | 552 | 7 |
| Всего, т/ км2 | 23,73 | 17,00 | 1,84 | 0,36 |
России с акцентом на средних и высших трофических уровнях. Для уточнения представлений о рыбопродуктивности были проведены комплексные бонитированные, т. е. оценочные съемки, включавшие около 30 экспедиций, объединявших ихтиологов, гидробиологов и океанологов. Бонитировкой был охвачен весь дальневосточный российский шельф, частично материковый склон и вся пелагиаль до глубины 1000 м. При тотальных подсчетах численности и биомассы рыб и беспозвоночных учитывались все, а не только промысловые гидро- бионты. В результате были получены достоверные данные о соотношении видов и групп в пелагических и донных сообществах (Шунтов, 1995).
Сначала съемки были проведены на шельфе. Оказалось, что в 80-е гг. биомасса донных рыб надалъневосточном шельфе от зал. Петра Великого до Берингова пролива составляла 3,8 млн т, из которых 50,5% приходилось на треску-’и навагу, 28,4% - на камбалу, 11,1% - на бычков и 10% - на прочих рыб. Наибольшие концентрации донных рыб были характерны для западнокамчатского шельфа - 23,73 т/км-, Карагинского и Олюторского заливов - 17,0, шельфа южных Курил - 13,45 т/км-1. В то же время на североохотоморском шельфе концентрации составляли только 1,84, в Анадырском заливе - 3,79, в Беринговом проливе - 0,36 т/км- (табл. 5.3).
Таким образом, несмотря на многолетний интенсивный промысел. запасы большинства рыб в камчатских водах оказались гораздо
выше, чем представлялось ранее. Это позволило более чем на 100 тыс. тувеличить квоты на вылов трески и камбалы. Съемками было опровергнуто существовавшее десятилетиями представление о том, что в результате перепромысла камбал на дальневосточном шельфе их ншлу заняли бычки и другие непромысловые рыбы.
Кроме того, новые данные подтвердили, что экологический потенциал многих массовых промысловых донных рыб весьма высок, и при регулировании рыболовства, которое началось с введения в 1977 г. Советским Союзом 200-мильной экономической зоны, резко ограничившей иностранный промысел в наших водах, их запасы начали возрастать.
Количество рыбы в том или ином районе в первую очередь.зависит от трофической емкости вод и условий воспроизводства. Для Мирового океана имеется лишь несколько примеров достоверных цифровых оценок экологической емкости конкретных регионов. Рассмотрим, к примеру, западнокамчатский шельф, наиболее богатый рыбой и промысловыми беспозвоночными район дальневосточных морей. Как известно, запасы донных гидробионтов, в том числе камбал и камчатского краба, сильно снизившиеся в 50-60-е гг. в результате интенсивного промысла, к началу 80-х гг. (благодаря введению экономической зоны) заметно возросли. Проведенный вто время комплекс исследований состава и структуры бентосного сообщества западнокамчатского шельфа показал, что продукция бентоса рыбами и хищными беспозвоночными явно недоиспользовалась. На основании этого был сделан вывод о возможности роста запасов донных рыб и краба на западнокамчатском шельфе.
Через несколько лет соотношение между продукцией бентоса и ее потребителями заметно изменилось, и к концу 80-х гг. потребление бентоса приблизилось к величинам его продукции (табл. 5.4). Если учесть, что не весь бентос является для рыб кормовым, в настоящее время кормовые ресурсы на западнокамчатском шельфе ограничивают рост донных рыб и крабов.
Бонитировочные съемки эгтипелагиали установили, что в ихтио- иенозах наибольшей части российской экономической зоны резко доминирует всего один вид - минтай. Например, в Беринговом море на его долю приходилось 80-90% от обшей массы рыб, в Охотском - 75-85%, только в Японском море, на юге Охотского, а также втихоо- кеанском южнокурильском районе в 80-е гг. доминировала сардина- иваси. В этот период в Беринговом и Охотском морях размножались высокочксленные стада минтая. Однако сработавший фактор плотности популяции, действие которого связано с ограниченностью экологической емкости среды, привел к быстрому сокращению минтая в Беринговом море к началу 90-х гг. В Охотском море численность
Биомасса и продукция (млн т) рыб и беспозвоночных па западнокамчатском шельфе
в 1980-с it.
(по: Шунтов, 1995)
минтая сохранялась на довольно высоком уровне влечение всей пер- вой половины 90-х гг.
Как отмечает В. П. Шунтов, до сих пор имеют1 место утверждения о подрыве запасов минтая в результате нерегулируемого промысла в этом районе. Однако в 80-90-е гг. потери минтая от хищников значительно превышали его вылов. Кроме того, сравнение суммарных потерь минтая от промысла и хищников с его продукцией (табл. 5.5) показывает, что крупномасштабное изъятие не превышало продукционных возможностей минтая.
Понять, почему Берингово и Охотское моря различались динамикой сокращения численности минтая, помогли гидробиологические съемки. В эти же 80-е гг. были получены огромные массивы информации по составу и ресурсам планктона во все сезоны и по всей плошали Дальневосточной экономической зоны России. Новые гидробиологические данные легли в основу расчетов продукции по трофическим уровням, а также построения схем биологического баланса конкретных морей.
Оказалось, что в 80-е гг. биомасса и продукция планктона и бентоса были заметно выше в Охотском, а продукция рыб и промысловых беспозвоночных - в Беринговом море. Это означало, во-первых, что в Беринговом море гораздо выше эффективность экосистем с позиций выхода продукции на высших трофических уровнях. Во-вторых, что здесь более четко работает фактор плотности, обусловленный внутри- и межвидовой конкуренцией за пищу (табл. 5.6).
Весьма существенно, что в Беринговом море значительна доля хищного планктона (табл. 5.7), особенно малоценных в кормовом
Среднегодовая продукция и потребление (млн т) мин тая в Охотском
и Беринговом морях в 1980-е гг.
(но: Шунтов, 1995)
отношении щетинкочелюстнътх (сагитт), в Охотском же - доля эвфа- узиид, излюбленной пищи многих рыб, в том числе минтая. Более сильное обострение трофических противоречий в Беринговом море подтверждает и увеличение числа альтернативных рыб, т. е. рыб, динамика численности которых находится в противофазе. Например, минтаю альтернативна сельдь: в период высокой численности минтая бывает мало сельди, и наоборот. В последние годы в пелагических сообществах происходит довольно быстрая замена теплолюбивых рыб на холодолюбивые. Закончилась эпоха сардины-иваси, значительноменьше стало минтая. В го же время возрастает численность альтернативных им более холодолюбивых видов японского анчоуса, сельди, мойвы и тихоокеанского кальмара. Поскольку новые объекты промысла пока не компенсируют убыль доминировавших в эпи- пелагиали. рыбопродуктивность дальневосточных морей по сравнению с 80-ми гг. снизилась.
Необходимо помнить, что в основе специфики сообществ обоих морей лежат различия в климато-океанологических условиях. Охотское море значительно суровее: низкие температуры оказывают сдерживающее влияние на воспроизводство многих рыб и промысловых беспозвоночных, хотя не ограничивают развитие планктона и бентоса. Очевидно, поэтому рыбопродукция в Охотском море отстает откор- мовой базы, т. е. биомассы и продукции планктона и бентоса.
Бонитировочные съемки 80-х гг. принесли новые данные по мелким мезопелэтическим рыбам. За исключением Японского моря, где их мало, по биомассе они (в основном миктофиды и серебрянка)
Продукция (г/м2) основных групп животных в Беринговом и Охотском морях
в 1980-с гг.
(по: Шунтов, 1995)
не уступают массовым эттипелагаческим рыбам - минтаю, сельди, лососям, мойве. В Беринговом море, например, при биомассе рыб, оцененной в 50-60 млн т, на долю мезопелагических приходилось 15 - 20 млн т В Охотском море это соотношение составляло 35 и 15 млн т. В тихоокеанских водах Камчатки и курильских водах мезопелагичес- кие рыбы вообще преобладают над всеми другими. Большая биомасса предопределяет их роль в сообществах, прежде всего в количестве потребляемого планктона. К сожалению, современная техника промышленного рыболовства не позволяет пока включить эти объекты в промысловую сферу.
Как уже отмечено, в начале 90-х гг. в экосистемах дальневосточных морей начались перестройки, связанные с наступлением более холодной эпохи. В океанологических работах этот факт еще пока не отражен, однако гидробиологи заметили его по динамике состава планктонных сообществ. Как можно видеть в табл. 5.7, в Беринговом и Охотском морях в 90-е гг. сильно уменьшилось количество самой массовой группы планктона - копепод. В то же время в Беринговом море возросла биомасса хищных сагитт, а в Охотском - эвфаузиид. Уменьшение количества копепод в Беринговом море является, очевидно, следствием усиления пресса со стороны хищного планктона, биомасса которого, превосходившая на порядок биомассу планкто-
Межгодовые изменения биомассы (пас. т) различных групп макронланкгона в Охотском и Бершновом морях в летний период
(по: Шунтов, 1995)
| Район | Год | Эвфау- зииды | Амфиподы | Копсподы | Щетинкоче люстные | Прочие |
| Южная часть | 19S6 | 24 050 | 4 190 | 60080 | 14 530 | 1850 |
| Охотске го моря | 1991 | 29 698 | 2 654 | 17 354 | 16 866 | 679 |
| | 1993 | 40 471 | 4 855 | 13 820 | 16 665 | 2 733 |
| Юго-западная | 1989 | 4 665 | 645 | 56 735 | 14 114 | 802 |
| часть Берингова | 1991 | 2891 | 457 | 6 985 | 20 777 | 2 374 |
| моря | 1993 | 6 131 | 1000 | 18281 | 39 201 | 2514 |
ноядных рыб, выросла еще больше из-за резкого сокращения рыбы, в первую очередь минтая. Увеличение эвфаузиид в Охотском море при сильном снижении массы копепод является, возможно, следствием пищевой конкуренции между различными группами планктона, тем более что для Охотского моря вообще характерно обилие эвфаузиид. Пелагические сообщества дальневосточных морей находятся в переходном состоянии: ‘‘ивасевый” период сменился на “сельдевый”.
Таким образом, биоценотическип подход к изучению биоресурсов дальневосточных морей позволил получить ясные представления о рыбопродуктивности дальневосточной экономической зоны. В 80-е и первой половине 90-х гг. были получены первые достоверные ряды наблюдений по динамике сообществ. Понимание необходимости иметь такие данные пришло сравнительно недавно. До этого преобладали так называемые объектные рыбохозяйственные исследования с акцентами на однофакторные связи конкретных видов и популяций с условиями среды, которые не позволяли иметь надежного прогноза промысла. Основной причиной недостаточности традиционных подходов при изучении ресурсов промысловых объектов является неучитывание состава биоценозов, в которых обитают промысловые объекты, т. е. игнорирование многообразных и сложных взаимоотношений каждого вида с его биотическим окружением. Трофические отношения являются основой функционирования природных сообществ, любое из которых предсташтяет собой пищевую сеть, по которой происходит перераспределение потоков вещества и энергии. Ясно, что при различных воздействиях извне объектами влияния будут не только отдельные популяции и виды, но и экосистемы в целом.
Как видим, рациональное природопользование и действительное управление биологическими ресурсами должны базироваться на экосистемных исследованиях, на знании ключевых связей в экосистемах и закономерностей их функционирования.
Поток энергии. Для осуществления любых жизненных процессов необходима энергия. Единственным источником энергии для зеленых растений является Солнце. В экосистемах, где солнечная энергия недоступна (например, дно океана), источником энергии для организмов служит окисление неорганических веществ.
Солнечная энергия, падающая на фотосинтезирующие органы растений, аккумулируется во вновь образующихся органических соединениях. Эта энергия используется продуцентами по-разному. Часть ее тратится на дыхание, т. е. на биологическое окисление (см. § 12, 13), часть запасается в виде вновь возникшей биомассы. Биомасса - это масса организмов определенной группы или сообщества в целом.
Некоторую долю созданной продуцентами биомассы съедают травоядные животные. Хищники потребляют травоядных животных и получают долю энергии. Часть энергии, полученной консументами с пищей, тратится на процессы, происходящие в клетках, а также выводится с продуктами жизнедеятельности в окружающую среду. Другая часть энергии идет на увеличение массы тела, рост и размножение и рассеивается в виде тепла.
Часть биомассы продуцентов, не съеденная животными, отмирает, и с отмершей биомассой аккумулированная в ней энергия поступает в почву в виде растительного опада.
Растительный опад, трупы и экскременты животных служат источником питательных веществ для редуцентов. Определенное количество энергии запасается в биомассе редуцентов, а часть рассеивается. Редуценты отмирают, и их клетки также разлагаются. Из продуктов разложения состоят органические вещества почвы.
Таким образом, энергия аккумулируется на уровне продуцентов, проходит через консументы и редуценты, входит в состав органических веществ почвы и рассеивается при разрушении ее разнообразных соединений.
Разобранный пример относится к наземным экосистемам. Подобным же образом происходят процессы и в водных экосистемах. Через любую экосистему проходит поток энергии, определенная часть которой используется каждым живым существом.
Цепи питания. Перенос энергии от ее источника (растений) через ряд организмов называют пищевой цепью.
Все живые организмы связаны между собой энергетическими отношениями, поскольку являются объектами питания других организмов. Травоядные животные (потребители первого порядка) поедают растения, первичные хищники (потребители второго порядка) поедают травоядных, вторичные хищники (потребители третьего порядка) поедают хищников помельче. Таким образом создаются пищевые цепи из продуцентов и консументов, которые на разных этапах, как это было показано в § 70, смыкаются с сообществом редуцентов (рис. 104).
Рис. 104. Пищевые цепи в наземных экосистемах
Пищевые цепи разделяют на два типа. Один тип пищевой цепи начинается с растений и идет к растительноядным животным и далее к хищникам. Это так называемая цепь выедания (пастбищная). Другой тип начинается от растительных и животных остатков, экскрементов животных и идет к мелким животным и микроорганизмам, которые ими питаются. В результате деятельности микроорганизмов образуется полуразложившаяся масса - детрит. Такую цепь называют цепью разложения (детритной).
На суше пищевые цепи первого типа состоят обычно из 3-5 звеньев, например: растения - овца - человек - трехзвенная цепь; растения - кузнечики - ящерицы - ястреб - четырехзвенная цепь; растения - кузнечики - лягушки - змеи - орел - пятизвенная цепь. Через пищевые цепи биогеоценозов суши подавляющее количество прироста растительной биомассы поступает через опад в цепи разложения.
В морях и океанах обнаружено несколько типов пищевых цепей. В морях, расположенных на шельфе (например, Баренцево море), это: фитопланктон - мелкие рачки - рыбы, питающиеся мелкими рачками (сельдь, мойва) - хищные рыбы (треска) - морские млекопитающие (гренландский тюлень); в открытых районах океана: фитопланктон - зоопланктон (инфузории, рачки) - кальмары и рыбы, питающиеся мелкими рачками (макрель) - хищные рыбы (тунец) - крупные акулы и дельфины. В редких случаях, например в зонах подъема к поверхности глубинных вод (зоны апвеллинга), пищевая цепь сокращается до трех основных звеньев: фитопланктон - мелкие рыбы (анчоус) - хищные рыбы. В водах Антарктики и некоторых других районах Мирового океана конечным звеном наравне с млекопитающими выступают хищные птицы. В районах открытого океана основная часть фитопланктона проходит через цепь выедания. На шельфе (до глубины в среднем 200 м), наоборот, большая часть биомассы, накопленной фитопланктоном, оседает на дно, где включается в детритную пищевую цепь (рис. 105).

Рис. 105. Пищевые цепи в океане
Все типы пищевых цепей всегда существуют в сообществе таким образом, что член одной цепи является также членом другой. Соединение цепей образует пищевую сеть экосистемы. Угнетение или разрушение любого звена экосистемы с неизбежностью отразится на экосистеме в целом. Поэтому вмешиваться в жизнь экосистем надо с большой осторожностью и осмотрительностью.
Экологическая пирамида. Пищевые сети внутри каждой экосистемы имеют хорошо выраженную структуру. Она характеризуется количеством и размером организмов на каждом уровне цепи питания. Как правило, при переходе с одного пищевого уровня на другой численность особей уменьшается, а их размер увеличивается. Например, в приведенной выше четырехзвенной цепи на 1 га травяной экосистемы насчитывается около 9 млн растений (первый пищевой уровень), свыше 700 тыс. растительноядных насекомых (второй уровень), больше 350 тыс. хищных насекомых и пауков (третий уровень) и всего три птицы (четвертый уровень). Как мы видим, образуется пирамида чисел, основание которой в 3 млн раз шире, чем вершина.
Только часть энергии, поступившей на определенный уровень биоценоза, передается организмам, находящимся на более высоком пищевом уровне. С уровня на уровень переходит около 10% энергии. Можно подсчитать, что энергия, которая доходит до пятого уровня (например, до орла в цепи: растения - кузнечики - лягушки - змеи - орел), составляет всего 0,01% энергии, поглощенной продуцентами. Таким образом, передача энергии с одного пищевого уровня на другой происходит с очень малым КПД. Это объясняет уменьшение числа и массы организмов на каждом последующем уровне и ограниченность количества звеньев в пищевой цепи.
Продукция экосистем. Любую экосистему характеризуют два важных параметра - биомасса и ее прирост за год, т. е. урожай. Прирост биомассы, созданной за единицу времени, называют продуктивностью экосистемы. Экосистемы суши имеют различную биомассу и продуктивность. Самой низкой биомассой растений и продуктивностью обладают тундры и пустыни, самой высокой - тропические дождевые леса. В тундре растениям не хватает тепла, в пустыне - воды. В лесном поясе тропиков много тепла и влаги. В открытом океане биомасса водорослей очень мала (продуктивность 1-2 т/га в год): их рост ограничен недостатком питательных элементов и света. Там, где света и питательных веществ много, например в прибрежных областях, биомасса значительно выше.
Несмотря на то что океан занимает 71% площади нашей планеты, его продуктивность в 3 раза, а биомасса водорослей в 10 тыс. раз меньше, чем продуктивность и биомасса растений суши.
Такая громадная разница в биомассе растений суши и океана объясняется следующей причиной. Основные продуценты суши - деревья, а океана - мелкие одноклеточные водоросли. Деревья растут медленно (низкий прирост), а живут долго, их биомасса накапливается за десятки и сотни лет. Водоросли в океане быстро размножаются. За год их поколения могут смениться десятки и сотни раз. Практически каждый день в океане создается масса водорослей, равная их запасу. Однако отмирание и оседание фитопланктона на дно вместе с поеданием его консументами быстро снижают массу одноклеточных водорослей. Устанавливается равновесие между прибылью и убылью продуцентов, и запас их постоянно остается низким.
Количественный учет потоков энергии и продуктивности биогеоценоза имеет большое практическое значение. Точный расчет потока энергии и продуктивности позволяет регулировать в экосистемах выход выгодной для человека биомассы живых организмов и представлять допустимые пределы ее изъятия.
Таким образом, рассмотрев поток энергии в биогеоценозах и цепи питания, мы увидели, что в сообществе живых организмов от звена к звену циркулируют основные питательные элементы и энергия. Автотрофы, аккумулируя солнечную энергию, потребляя углекислый газ и элементы минерального питания, создают органические вещества, которые служат пищей гетеротрофам. Гетеротрофы, разрушая органические вещества, обеспечивают себя энергией и освобождают элементы питания для автотрофов. В циркуляцию веществ и энергии включены не только живые организмы, но и среда их обитания.
- Откуда организмы получают энергию и как они ее расходуют?
- Как связаны в потоке энергии продуценты и консументы?
- Что является источником энергии для редуцентов?
- В чем разница между продуцентами суши и океана?
- Приведите пример экологической системы своей местности и охарактеризуйте ее.
- В средних широтах приток солнечной энергии за год 3,8х10 10 кДж/га. Гектар леса производит за год 10 тыс. кг древесины и листьев. В каждом грамме производимых веществ заключено в среднем 19 кДж. Сколько процентов падающей энергии использует лес?
Гренландский тюлень — удивительное животное. О его особенностях, повадках, местах обитания мы расскажем в этой статье. По-другому это млекопитающее еще называют лысун.
Где живет этот зверь?
Ареал обитания этого представителя фауны достаточно обширный, он распространен в арктических водах. Встречается гренландский тюлень в в Белом море, на побережье и Представителя этого вида можно встретить севернее острова Ян-Майен. Вне брачного периода животные занимают и другие территориальные просторы, например, в Баренцевом, Также этот вид можно встретить даже в атлантических уголках Канады и острова Гренландия.
Особенности
Гренландский тюлень — самый многочисленный вид семейства под названием «настоящие тюлени». Несмотря на его широкое разнообразие, лысуна очень легко отличить от других представителей «настоящих тюленей».
Первое и самое главное, что следует отметить — это специфический и уникальный окрас, который имеет этот арктический житель. При рождении детеныши гренландского тюленя обладают зеленоватым цветом шерсти. По истечении нескольких деньков цвет шерсти малыша и сама её структура изменяется. Она становится полой и прозрачной. Солнечные лучи сквозь такие полые ворсинки легко попадают на черное тело и прогревают кожу. В среде обитания гренландского тюленя это как никогда кстати.
Когда малыш вырастает, становится самостоятельным и отказывается от молока, у него проявляются все особенности, которыми обладает гренландский тюлень, фото явно показывает главный отличительный признак — это ярко выраженные полоски с двух сторон по спине. У самцов он выражен сильнее, чем у самок этого вида. Форма полосок напоминает полумесяц, цвет - темно-коричневый. Притом что цвет шерсти тюленя — серый. Полоски соединяются в районе крестца в верхней части спины. Кстати, голова тоже имеет отличный от основного оттенка цвет — бурый, это еще одна отличительная особенность такого зверя, как гренландский тюлень, фото которого вы видите в нашей статье.
Размеры
Второе, что хотелось бы отметить — это достаточно большие размеры такого северного представителя. Длина гренландского тюленя составляет не менее 180 сантиметров, оптимальные размеры - от 180 до 185 см. Конечно, попадаются как более крупные особи до 190 см, так и сравнительно небольшие — 160 сантиметров.

У многих представителей животного мира существуют резкие перепады в размерах самцов и самок. В случае рассматриваемого вида разницы практически никакой нет. Самки этого тюленя могут быть не намного меньше самцов. Вес этих зверей колеблется от 140 до 160 килограмм.
Большую долю в формировании массы тела составляет значительная жировая прослойка. Толстый слой жира — это своеобразный залог здоровья. Он позволяет выдерживать невыносимо низкие температуры, которые не редкость для арктических вод. Кроме того, жир придает наиболее совершенную обтекаемость телу этих зверей. А этот факт прямо пропорционально влияет на скорость передвижения животного в воде и прочие параметры в плавании.
Как мигрирует тюлень гренландский?
Тип миграции этого вида мы будем рассматривать далее. Сейчас отметим тот факт, что гренландский тюлень практически всегда находится в процессе переселения, он постоянно передвигается вдоль кромки арктического льда. В статическом — относительно неподвижном положении - он находится только при одном из трех условий: в процессе спаривания, в период произведения на свет детенышей, а также во время линьки.

Миграция таких животных связана с переселением из открытого моря к стационарным лежбищам. Самки этого представителя фауны практически до самого момента рождения малышей плавают в арктических водах. Роды у гренландского тюленя происходят буквально в первых числах марта. Место для произведения на свет потомства должно быть просторным, прочным и с толстым снежным покровом.
После родов самки сбиваются в особые стаи, подобные «яслям», только они огромные по размерам. Наиболее научное название — щенные залежки. Вот в таких условиях и появляется гренландский тюлень, интересные факты об этом животном мы поведаем вам немного позже. Так вот, первую неделю жизни самка лежит вместе с детенышем, позже, через 7-10 дней, она начинает выходить в открытое море и основное время проводит в воде, на лед выходит только для молочного кормления.
Первый — двойня рождается очень редко, в основном на свет производится по одному детенышу. Размеры его таковы: длина - 1 метр, вес - примерно 8 килограмм. После рождения малыш гренландского тюленя в день набирает порядка 2 килограмм. Основные питательные вещества передаются с молоком матери, которое очень жирное, собственно, это и способствует быстрому набору веса.

Еще один интересный факт связан с линькой лысуна. Линька этого безобидного животного - достаточно продолжительный и болезненный процесс, происходит который в конце марта. Кроме волосяного покрова, обновляется и сам кожный покров. Во время линьки животное сильно теряет в весе, потому что практически не ест. По завершении этого действа и с возобновлением активного образа жизни вес быстро восстанавливается.
Важно знать!
Гренландский тюлень — это удивительное животное, безобидное, которое охотится только на рыбу, водящуюся в местах его обитания. Врагов у жителя арктических холодных вод практически нет, основные млекопитающие, которые представляют опасность, — это белые медведи и касатки. Но для первых достаточно тяжело поймать на льдинах тюленя из-за его маневренности. Касатки же редко представляют опасность для этих зверей из-за своего Обычно когда в воды приплывают касатки, тюлени уже мигрируют в более безопасное место.

Главную опасность для таких зверей представляет человек — это безоговорочно и несомненно. Урон популяции гренландских тюленей человек наносит как прямо, так и косвенно. Такие животные истребляются непосредственно ради ценного жира. Но этот промысел ограничен квотами, позволяющими сохранить популяцию. Гораздо страшнее оказывается чрезмерная добыча рыбы. Ведь именно она является основным рационом для лысунов, кстати, и не только для них.
Теперь вы знаете, кто такой гренландский тюлень, чем он питается и где обитает. Кроме этого, мы поведали вам несколько интересных фактов.
Эта статья посвящена гренландскому тюленю. Она расскажет о том, где обитает, чем питается, как передвигается на суше и под водой это животное, а также о том, кто такие бельки, зеленцы и серки. Приведено описание гренландского тюленя и интересные факты из его жизни.
Иначе называемый лысуном данный вид морских млекопитающих весьма распространен в арктических водах. Зоологи это животное относят к семейству настоящих (или истинных) тюленей.
Главное отличие гренландского тюленя от прочих представителей группы ластоногих (а это моржи, морские львы и котики) в том, что они не имеют ушной раковины. Вместо них по бокам головы у них лишь маленькие отверстия.
Описание
Наибольшая длина тела лысуна достигает двух метров (обычная - 180 см) при массе более 150 центнеров.
Характерный окрас не позволит спутать это животное с другими представителями вида. Основной цвет шерсти - серебристо-серый, но голова черная, а в области плеч с обеих сторон проходят черные же полосы. В целом этот узор напоминает арфу, вот почему по-английски этот вид называют Harp seal, что означает арфа-тюлень. Впрочем, самки не обладают столь заметным украшением шкуры - их "арфа" может распадаться на отдельные пятна и окрас иметь побледнее.
Под кожей у гренландского тюленя находится толстый слой жира (ворвани), среди прочих играющего роль теплоизоляционного слоя, который позволяет животным легко переносить холод арктических вод.
Эти существа обладают прекрасным зрением и отличным обонянием. Под водой они способны развивать вполне приличную скорость, что помогает им в охоте.
Продолжительность жизни гренландского тюленя в дикой природе составляет не более 20 лет.
Ареал
Лысуны, как и прочие млекопитающие, относящиеся к этой группе, живут в арктических морских водах. Обычно они предпочитают держаться у кромки льда, особенно если это льды дрейфующие. Для размножения или просто отдыха животные выползают на лед.
На вопрос о том, где обитает гренландский тюлень, ответить не так уж и просто. Ученым известны три отдельные популяции лысунов в зависимости от места обитания и размножения. Что это за зависимость? Дело в том, что все тюлени для продолжения рода мигрируют в районы, находящиеся южнее.

Например, живущие в обычный период в Баренцевом и Карском морях в брачный мигрируют в Белое море. Там, на больших льдинах они образуют "щенные залежки", где самки могут охотиться и выкармливать своих детенышей.
Другая популяция этих тюленей обитает у канадских и гренландских побережий Атлантического океана, а брачный период проходит для них у Лабрадора и Ньюфаунленда, в заливе Святого Лаврентия.
Наконец, еще одна группа особей данного вида водится вблизи норвежского архипелага Шпицберген и берегов восточной Гренландии. "Играть свадьбы" они отправляются к острову Ян-Майен, находящемуся между Гренландским и Норвежским морями.
Пища для гренландского тюленя
Все ластоногие плотоядны. Зоологи указывают на их родство с различными псовыми - собаками, лисами, волками, койотами и даже медведями.

Взрослые тюлени обычно держатся вместе и охотиться предпочитают сообща. А чем питается гренландский тюлень? Летом пищей для него являются рыба, ракообразные и моллюски. Среди последних предпочитаемыми будут кальмары, осьминоги, лобстеры. В зимний период рацион почти полностью состоит из сельди, мойвы, морского окуня и др.
Плавает и ползает
Природа создала тюленя животным, обитающим в основном в водной стихии. Его тело веретенообразной формы с ластами и хвостом приспособлено для быстрого передвижения под водой. Ворвань также способствует лучшей обтекаемости тела.
Перед тем как нырнуть (а глубина погружения в воду тюленя может составлять порядка 200 метров), это животное совершает серию учащенных вдохов, стремясь наполнить кровь кислородом, но перед погружением оставляет в легких лишь минимум воздуха, чтобы тот не вытолкнул его из воды и не помешал охоте. Под водой морской охотник способен находиться от 5 до 15 минут.
Как же передвигается гренландский тюлень по суше? Дело в том, что у всех животных этого вида и истинных тюленей вообще задние ласты не могут сгибаться и подворачиваться так, чтобы можно было опираться на них при ходьбе. Для того чтобы переместиться с места на место по суше, они вынуждены совершать волнообразные движения туловищем.
Брачный период и рождение щенков
Как уже говорилось, для продолжения рода гренландские тюлени откочевывают в районы, находящиеся южнее, где сбиваются большими колониями на льдинах. В этой колонии могут одновременно находиться порядка десяти тысяч особей. Здесь самцы, используя свои клыки и ласты, устраивают с соперниками брачные бои, борясь за дам.

Приблизительно через 11 месяцев у каждой из самок рождается по тюлененку. В этот период самки сбиваются в свою "женскую" колонию и выкармливают детеныша самостоятельно, отдельно от самцов. А те, удалившись от забот воспитания подрастающего поколения, живут также на другой льдине, охотясь и отдыхая.
Самки тюленей хотя и обитают поблизости друг от друга, не принимают чужих щенков, кормя только своего и отталкивая чужаков. Но вернувшаяся с охоты мамаша легко находит своего тюлененка, где бы он ни находился, по запаху.
Какой он, детеныш тюленя?
Первые дни из-за влияния околоплодных вод шерсть малыша белая со слегка зеленоватым оттенком. Это зеленец. Новорожденный питается молоком матери, которое в 12 раз жирнее коровьего, и быстро растет. Ежедневно он прибавляет в весе по паре килограмм.
Постепенно шерсть его очищается и становится белого цвета. Этого тюлененка называют уже белек. Природа позаботилась о том, чтобы маленький тюлень был малозаметен на снегу для хищников - ведь ни плавать, ни защитить себя самостоятельно он еще не может.

Регуляция тепла в его теле происходит за счет постоянного дрожания. Кроме того, волоски шерсти бельков, полые внутри, почти прозрачны и пропускают солнечные лучи к самой коже.
Через 12 дней после рождения самка перестает кормить тюлененка и вообще забывает о нем. Еще через десять дней, исчерпав свои жировые запасы, накопленные ранее, проголодавшийся белек переживает первую линьку и становится серком, настоящим молодым тюленем с характерным для своего вида окрасом. Теперь мех его не намокает при погружении в ледяную воду, и он может самостоятельно охотиться.
Океан подо льдом
Тюлени и пингвины, живущие во льдах, рыбы из семейства нототениевых и все уникальные жители полярных морей обитают в среде, мало известной человеку. Установлено, что из глубоководных беспозвоночных более всего в полярных морях распространены кальмары, но многие их виды до сих пор знакомы нам лишь по клювам, найденным в желудках тюленей и китов!
Чтобы понять море и заключенную в нем жизнь, надо заглянуть в него по вертикали, сквозь все пласты, образующие сложные, постоянно изменяющиеся слои моря. Каждый слой переходит в последующий постепенно. Различаются они в основном плотностью, или удельным весом, воды, который зависит прежде всего от ее солености и температуры. Чем солонее вода, тем она тяжелее; чем выше ее температура, тем она легче. Поэтому более теплая и пресная вода обычно скапливается у поверхности, а более соленая и холодная опускается на дно. И в каждой такой водной массе, как принято называть эти слои, живут сообщества организмов, проспособившихся к ее уровню солености, температуре и постоянному мраку, царящему на глубине нескольких сотен метров. Итак, хотя в пелагиали морей и океанов нет ни стен, ни потолков между слоями, они четко отделены друг от друга свойствами самой воды.

Основа основ жизни в полярных океанах



В холодную и темную полярную зиму первичная продукция фитопланктона под морским льдом и в полярных водах крайне бедна. Естественно, что зависящие от фитопланктона живые существа - зоопланктон - в свою очередь не активны, т. е. не продуктивны. Между тем, зоопланктон служит основной пищей организмам, обитающим в этих морях. В зоопланктоне преобладают ракообразные, такие, как веслоногие рачки Calanus (на севере) или южная Euphausia, похожая на креветку (на юге). Оба эти вида зимуют на больших глубинах, и лишь когда подо льдом или в толще воды зацветает фитопланктон, поднимаются вверх. У многих веслоногих ракообразных обмен веществ протекает достаточно активно только при изобилии фитопланктона, а оно длится месяц, самое большее два. Зимой, в бездеятельном состоянии, они дрейфуют на глубине в одном направлении, а находясь во время и после весенне-летнего кормления на поверхности моря - в другом, и таким образом остаются в пределах узкой зоны, ограниченной в широтном направлении. Примечательно, что различные виды антарктических ракообразных, в частности криль, придерживаются в Южном океане обособленных участков, соответствующих определенным.


От этого в свою очередь зависит широтам, распространение охотящихся на них животных. Из-за кратковременности сезонного питания некоторые зоопланктеры размножаются только один раз в два года. Взрослые особи производят обычно большие яйца с желтком, из которых к моменту весеннего цветения вылупляются крупные молодые рачки. И взрослые, и молодь накапливают высококалорийный жир, который позволяет им пережить зиму. Возможно, на Земле нет второй столь стройной экосистемы, как та, что существует в Южном океане. Ее северная граница проходит по антарктической конвергенции, главную роль в ней играют ракообразные - эвфаузииды, известные под названием криль (Euphausia superba). Они служат главным источником пищи для многих видов, населяющих Южный океан. Ученые только недавно начали проникать в тайны его экосистемы и пришли к выводу, что ее главной движущей пружиной, без сомнения, является динамика криля. В наибольшем изобилии он встречается там, где богатство питательными веществами сочетается с другими факторами, благоприятствующими продукции основной пищи криля -фитопланктона, а такие условия складываются прежде всего в Южном океане, к югу от Атлантики. Криль питается, выцеживая фитопланктон из воды с помощью «фильтровальной корзины», образуемой вытянутыми вперед, вниз и в стороны ловчими ногами. Летом в верхнем стометровом слое на открытых участках и среди дрейфующих льдов кишит криль. По данным последных исследований, биомасса таких скоплений достигает иногда 10 млн. т, что равнозначно по массе почти двум миллионам слонов. Распределение в океане животных, питающихся крилем - рыб, пингвинов, тюленей, китов, -безусловно зависит от наличия этих огромных запасов пищи. Подсчитано, что постоянный запас криля в Южном океане составляет не менее одной трети миллиарда тонн. Из них тюлени-крабоеды (Lobodon carcinophagus) поедают около 100 млн. т. До начала промышленной добычи китов, когда их было намного больше, чем в настоящее время, они были главными потребителями криля, возможно даже опережая крабоедов. По мере избиения китов возрастала численность тюленей-крабоедов, пингвинов, питающихся крилем, кергеленского котика (Arctocephalus australis) и, может быть, некоторых видов рыб. Последние, впрочем, потребляют иные виды криля и в иных количествах. Китам требуется ежегодно количество криля, лишь в 4 раза превышающее вес их тела, крабоедам - в 20-25 раз, пингвинам - в 70 раз. Это объясняется отношением поверхности тела к его массе: чем больше это отношение, тем больше расход пищи на теплоотдачу. Вот и получается, что самое крупное на Земле животное - синий кит весом в 100 т уничтожает в год 400 т криля, а 100 т пингвинов съедают за это же время 7000 т! По-видимому, истребление китов человеком привело к тому, что та пища, которой прежде кормились киты, теперь достается тюленям и пингвинам, чья биомасса значительно меньше. В настоящее время в экосистеме Южного океана на каждого кита приходится по весу в 5 раз меньше тюленей и в 25 раз меньше пингвинов - весьма малоэффективное использование продукционных процессов Южного океана!

Это вовсе не означает, что самой экосистеме Южного океана угрожает опасность. Скорее следует предположить, что в результате происшедших изменений сложилось новое „статус-кво", при котором прежнее изобилие гигантских китов уже никогда не вернется. Этому мешает ожесточенная конкуренция пришедших на смену популяций животных, которые размножаются быстрее. Но раз Южный океан и его обитатели находятся в столь сильной зависимости от криля, возникает другой вопрос: что произойдет, если его количество уменьшится на порядок? Установится ли новое равновесие или удар по доминантному виду подорвет его навсегда? Ученые пока далеки от того, чтобы ответить на этот вопрос.



Хохлач, чтобы удержать других самцов от посягательств на свою территорию, принимает демонстративно угрожающую позу: раздувает эластичную перегородку между своими носовыми отверстиями и выпячивает ее наружу. Попеременно опуская и поднимая голову, тюлень выталкивает "воздушный шар" то из одной, то из другой ноздри
На морском льду

Животные, которым постоянным субстратом служит морской лед, некогда жили на суше. Морские млекопитающие произошли от наземных. Они, приспособились к жизни в море, тело их постепенно приобрело очертания, напоминающие рыб, хотя они сохранили многие характерные особенности млекопитающих. Морские котики и сивучи, например, продолжают щениться на суше, моржи время от времени выходят на землю. Зато другие морские млекопитающие, такие, как настоящие тюлени (Phocidae) Арктики и Антарктики, редко изменяют морскому льду, на нем они щенятся, кормят детенышей, отдыхают. Киты приспособились к морю не хуже тюленей, но в отличие от них не используют лед для какой-либо определенной цели, разве что как временное укрытие. Морской лед образуется из морской воды и тянется на сотни километров вдали от суши, он является неотъемлемой частью моря, и тех зверей, которые отдыхают или живут на нем, по праву следует считать морскими в полном смысле этого слова. Морской лед играет важнейшую роль в жизни тюленей и моржей, а также некоторых видов у гренландских китов, никогда не покидающих ледяную обитель.


Морской лед занимает важное место в жизни тюленей и моржей, а также некоторых видов китов. От него зависит существование сообщества в течение всего года, а следовательно и его социальная структура. В Антарктике на льду обитают четыре вида тюленей: Уэдделла, крабоед, Росса, морской леопард, в Арктике - шесть: кольчатая нерпа, крылатка, лахтак, ларга, лысун, хохлач. Все они относятся к настоящим тюленям и сильно отличаются от сивучей и морских котиков семейства Otariidae, имеющих заостренные наружные ушные раковины небольшой величины. Представители семейства Otariidae довольно хорошо ходят, даже, можно сказать, бегают по земле на всех четырех. Phocidae, лишены наружных ушных раковин, вместо них по обеим сторонам головы расположены лишь маленькие слуховые отверстия, что свидетельствует об очень тонком слухе. Тело их имеет веретенообразную форму, они утратили способность при ходьбе подтягивать задние конечности в помощь передним. В воде передвигаются колебательными движениями задних ластов вправо и влево, а по твердой поверхности - гусеничным ходом. При этом далеко не все виды тюленей тихоходны: крабоед, крылатка и некоторые другие скользят, подобно пресмыкающимся, по гладкой поверхности льда со скоростью бегущего человека. Тем не менее вне водной среды двигательные возможности тюленей ограничены, и им трудно спасаться бегством от наземных хищников. В Антарктике, где хищные звери отсутствуют, тюлени почти не боятся людей, приближающихся по льду. Иное дело Арктика: там на берегу ластоногих подстерегают охотники-эскимосы, белые и бурые медведи, лисы, и тюлени, чрезвычайно осторожные, никого к себе не подпускают.



Моржи, обитающие только в северных и арктических морях, представляют собой нечто среднее между тюленем и сивучем. Наружные ушные раковины у них отсутствуют, но с помощью задних ластов они передвигаются, хотя и неуклюже, по земле и по морскому льду. В воде они ведут себя в основном как настоящие тюлени, но иногда, уподобляясь сивучам, пускают в ход передние ласты и плывут, взмахивая ими, словно крыльями. Тюлени десяти видов и моржи, обитающие во льдах, активно ищут морской лед - на нем они отдыхают. Большинство китов, напротив, избегает слошных ледяных полей. Только три их вида проводят в ледяных водах круглый год: гренландский кит, нарвал и белуха. Остальные киты, в частности полосатики (к ним относятся гигантские синие киты и близкие виды), плавают в морях с ледяным покровом много времени, но все же не постоянно. Хотя они посещают полярные угодья, чтобы кормиться, их адаптация ко льду не столь совершенна, как у гренландских китов, никогда не покидающих ледяную обитель.
Морской лед занимает важное место в жизни морских млекопитающих на всех ее этапах. Тюлени, моржи, одним словом все обитатели моря кроме китов, вынуждены выходить на лед, чтобы отдыхать, размножаться, выкармливать детенышей. Каждый вид этих животных приспособился не только к различной пище, но и к различным типам льда, и в результате у каждого сложилась своеобразная социальная структура. Моржи и тюлени Уэдделла общительны до предела и держатся всегда большими группами. Лахтаки предпочитают одиночество, чувство территориальности у них развито очень сильно, они занимают участок льда в единоличное владение и защищают его от посягательств со стороны. Другие тюлени соединяются в пары и используют льдины для спаривания, и, наконец, есть такие, которые заимствуют понемногу от каждого образа жизни.


Социальная жизнь моржей
Наши познания о морских млекопитающих ограничиваются в основном тем, что мы видим на поверхности моря. Их социальная жизнь предстает нам наподобие айсберга, у которого видна только его верхняя часть. Как ведут себя тюлени под водой - мы не знаем, остается только догадываться по отдельным намекам, а это не идет ни в какое сравнение с наблюдениями, скажем, над птицами.
Убедительный тому пример - моржи. Эти коммуникабельные животные на протяжении всего года проявляют величайшую общительность. Более того, они живут чуть ли не в обнимку со своими соседями, непроизвольно прижимаясь к ним. Это отличает их от многих тюленей, которые, тоже будучи любителями общества, собираются в большие стада, но избегают физических контактов. Лежбище моржей выглядит со стороны единой коричневой массой, хотя его социальная структура представляет собой сложную иерархическую лестницу с неукоснительным соблюдением степеней социального превосходства самцами, самками и молодняком. Вероятно, моржи не только входят в стада с их социальной иерархией, но и объединяются в отдельные группы, которые могут действовать вполне автономно. В мае каждого года на морском льду собирается залежка, состоящая из самок, молодняка и нескольких зрелых самцов. Несколько в стороне иногда располагается группа беременных моржих, близких к разрешению от бремени. Залежка вместе со всеми подгруппами может насчитывать от нескольких десятков до нескольких тысяч особей.

Детеныши появляются на свет, когда ледяной покров начинает таять под лучами весеннего и летнего солнца. Его граница соответственно отступает на север, а вслед за ней передвигается и все стадо. На север вместе со льдом уходят независимо от самок и группы, состоящие из одних самцов, но лишь немногие из них перебираются вместе с самками и молодняком в Чукотское море, на общие с ними льдины. Многие моржи отделяются от главного стада и выходят на острова у русского или американского берега Берингова пролива. Здесь, на суше, их сравнительно легко наблюдать. Социальное превосходство среди самцов устанавливается в зависимости от размеров их туловища и бивней: чем крупнее морж, чем длинее его бивни, тем выше он стоит над другими животными. Это и определяет иерархию, сильно напоминающую хорошо известную очередность цыплят у кормушки, что очень удобно, ибо избавляет от необходимости вести кровопролитные битвы. Правда, столкновения все же происходят - о них свидетельствуют шрамы на шеях животных, но стоит моржу продемострировать свои могучие бивни, и его противник, с бивнями покороче, идет на попятный. Бивнями вооружены и самки, но пока не удалось окончательно выяснить, устанавливается ли с их помощью такая же иерархия, что и у самцов, хотя это подтверждается отдельными наблюдениями. Изменения в ледовой обстановке, происходящие осенью и в начале зимы, заставляют самцов покидать летние резиденции. В январе в центральной и южной частях Берингова моря они встречаются со стадами самок, которых распространение льда оттеснило на юг. К этому времени у самцов начинается гон, у самок -течка. Моржи достигают полового созревания позднее всех млекопитающих: самки могут зачать лишь в возрасте десяти лет, самцы готовы к совокуплению не раньше пятнадцати. Кормление детеныша молоком длится почти два года, поэтому моржиха щенится через два, а то и три года. В конце января моржи начинают ухаживать за своими избранницами (это замечательное явление описано ниже). Стада собираются на прочном ледяном покрове толщиною не менее одного метра, далеко от его кромки, где не ощущается волнение моря. Особенно им нравятся такие участки, где ледяной покров разорван на большие ледяные поля, между которыми простираются реки-разводья. Одна треть самок к этому времени уже беременна, другая треть вскоре должна родить, остальные еще не покрыты.

Бивни, иногда достигающие у взрослого самца метровой длины, служат ему для установления превосходства над другими самцами, а иногда помогают вылезать из воды. Вопреки установившимся представлениям, морж не пользуется ими для выкапывания моллюсков со дна. А вот " бакенбарды" "-вибриссы облегчают ему поиски пищи на донном грунте

Никому пока не удалось наблюдать и описать роды у моржей, но если судить по сообщениям о тюленях, то они должны происходить быстро. Новорожденный и мать безусловно обмениваются какими-то звуками, скорее всего моржиха тихо ворчит, а детеныш издает выразительные возгласы "о-о-окс?". Возможно, они также дышат «рот в рот» - поочередно вдыхают и выдыхают друг в друга. Так они знакомятся. На детеныше мало жира, весит он при рождении всего лишь 50 - 70 кг. Шкура светло-серая. Он умеет плавать, но пока еще плохо.
После появления на свет детеныш не сразу входит в воду. Он остается на льду, рядом с матерью, и старается держаться с подветренной стороны. Таковой обычно оказывается брюхо матери с четырьмя сосками, из которых поступает молоко, по жирности превосходящее коровьи сливки. В нем 35 процентов жиров, 6 процентов белков, сахар отсутствует. Как показывает поведение животных в неволе, детеныш сосет понемногу, но часто, вероятно от шести до десяти раз в день. Вскоре детеныш начинает реже прикладываться к соскам, но зато всякий раз высасывает больше молока.
Съедаемое моржонком за день тянет, вероятно, одну десятую его собственного веса, мать же теряет раза в два больше, пока он не начинает плавать: в это время она и сама может кормиться. Несмотря на огромное количество потребляемой пищи, моржи растут далеко не так быстро, как тюлени. Малыш чаще всего года два не покидает мать. Сосет он все реже, но она защищает его, учит, согревает. К концу первого месяца жизни моржонок накапливает столько жира, что холод ему уже не страшен, и он часто и подолгу плавает. Первые месяцы его жизни обычно протекают гладко. Погода стоит мягкая, даже на дальнем севере Чукотского моря, где собираются стада почти из одних самок. Солнце стоит высоко, лед тает. Но в начале сентября море опять замерзает. Стадам приходится перекочевывать на юг, намного опережая нарастающую кромку льда. К осени молодой морж уже умеет хорошо нырять и питается двустворчатыми моллюсками и другими придонными беспозвоночными. Ему требуется много сил, так как при отступлении на юг часто приходится целыми днями плыть в ледяной воде. К концу первого года жизни это уже сильное животное, весящее намного больше 200 кг. За год он высосал свыше 2 000 л молока и съел больше тонны моллюсков и других беспозвоночных.
Песни под водой
Одно из наиболее изученных морских млекопитающих - по его поведению как на воде, так и под водой - тюлень Уэдделла в Антарктике. Это очень мирное животное близко подпускает к себе людей и переходит к обороне только в случае крайней нужды. Обитает он на припае, следовательно, отдельные его популяции легко доступны с антарктических научно-исследовательских станций. Кроме того, можно нырять вместе с этими тюленями и наблюдать их поведение подо льдом. Такие опыты проводились в середине 60-х годов и показали, что вблизи своих лежбищ и самцы, и самки производят множество шумов, даже издают нечто вроде трели наподобие продолжительного чириканья механической канарейки. Звуки эти тюлени издают, очевидно, только весной, в брачный период. Причем впечатление такое, что одни тюлени выводят трели, а другие к ним прислушиваются. А поскольку эти «песни» раздаются чаще всего около продушин во льду, логично предположить, что таким образом тюлени ограждают территорию вокруг своей продушины. Возможно также, что самцы трелями защищают выбранную брачную территорию, а самки - участок льда вокруг детенышей. Не исключено, что дыхательных отверстий во льду не хватает для всей популяции. В таком случае песня должна дать понять другим тюленям: «Это мои владения». Никто никогда не видел, чтобы дикий морж распевал под водой, но с помощью подводного микрофона - гидрофона можно услышать его пение. И какое! Начинается оно с двух коротких, словно удары, быстро следующих друг за другом звуков «тэп-тэп-буоу...инг...г...г...г», которые замыкает как бы звон колокольчика. Эта музыкальная фраза исполняется до четырех раз, затем слышатся многократные повторения «тэп-тэп», завершающиеся кодой, точно как в присказке: «Бриться-стричься-два гроша» ( Чрезвычайно популярная в Америке присказка "Shave-and-a-haircut-two-bits" произносится с непременным повышением тональности в конце фразы (на словах "two bits"). - Прим, перев. ). И снова «тэп-тэп-тэп», и, может быть, его опять венчает «инг-г-г-г», после чего морж выныривает на поверхность воды, испуская сквозь губы толщиною с хороший бифштекс короткий тихий свист. Так и звучит «песня» без начала и без конца, но как на нее реагируют самки -неизвестно, ведь никому не посчастливилось стать свидетелем спаривания моржей. Она распространяется под водой на несколько километров, возможно, ее назначение - оповещать о территориальных правах самцов или держать соперников на расстоянии, а может, сообщать самкам, что будущие супруги готовы к спариванию. Житель Арктики морской заяц, или лахтак (Erignathus barbatus), также принадлежит к «певцам». Тюлени этого вида выбирают себе для жилья довольно толстый паковый лед. Весной гидрофон улавливает издаваемые ими громкие трели по нисходящей гамме, напоминающие пение тюленей Уэдделла, но, пожалуй, даже более музыкальные, даже с модуляцией тональностей, причем одна вибрирующая мелодия накладывается на другую.
Тюлени и моржи - не единственные в полярных районах морские млекопитающие, чьи голоса раздаются под водой. Белого кита, или белуху (Delpinapterus leucas), за его вокальные данные прозвали морской канарейкой. Нарвал (Monodon monoceros) производит странный шум наподобие скрипа. Гренландский кит (Balaena mysticetus) ворчит басом. А вокруг стада лысунов (Pagophoca groenladica), когда они весной собираются для спаривания и деторождения, шум стоит, как около скотного двора, полного свиней и кур. По-видимому, в полярных морях звук является действенным средством связи и раздела территорий. Распространяется он в воде со скоростью свыше полутора километров в секунду, т. е. в пять раз быстрее, чем в воздухе, и на большие расстояния. Установлено, что многие морские млекопитающие могут сообщаться между собой даже на расстоянии 10 - 20 км. Иными словами, входящие в одно стадо животные могут рассеяться на площади 300 кв. км и тем не менее поддерживать связь друг с другом. Теоретически возможно, что некоторые крупные гладкие киты способны входить в контакт на расстоянии свыше 100 км, так как они производят очень низкие звуки, которые распространяются намного дальше, чем высокие. Если это действительно так, то эти киты поют, т. е. поддерживают социальные контакты, на территории свыше 30 тыс. кв. км! Но это, конечно, всего лишь предположения.
Жизнь китов
Гренландский, или полярный, кит - пожалуй, самый необычный из гладких китов: он единственный из них круглый год проводит в полярных морях и не мигрирует в более теплые края. Эти киты встречаются в водах Канадского архипелага и вблизи Гренландии, но больше всего распространены в Беринговом море. Определить их численность на сегодня более или менее точно трудно, но скорее всего в настоящее время она не превышает одной десятой части прежней численности. Приводить какие-либо числовые показатели - дело весьма рискованное, ведь никто еще не придумал способ подсчета китов с точностью свыше 50 процентов. Зимой гренландские киты, обитающие в Беринговом море, распространяются по его центральной части, имеющей прочный ледяной покров, но все же не настолько прочный, чтобы кит не мог набрать воздуха у поверхности воды. В этом ему помогает расположенная на голове выпуклость с ноздрями, так называемыми дыхалами. Ею кит проламывает полуметровый и даже более толстый лед. Чтобы вдохнуть воздуха, ему достаточно трещины во льду или узкой полыньи между ледяными полями. Когда над нею внезапно возникает V-образная двойная петля конденсировавшегося дыханья - это говорит о том, что кит всплыл. Вскоре после того как кит набирает свежего воздуха и опускается под лед, петля рассеивается. Гренландские киты приступают к миграции весной, когда небольшими группами собираются в районе довольно тяжелых льдов. Они намного опережают тюленей и моржей: Берингов пролив проходят в начале мая, тогда как моржи поспевают туда двумя неделями позднее, а некоторые тюлени только после середины июня. К середине мая гренландские киты уже находятся далеко в Чукотском море, в июне входят в море Бофорта, своих летних угодий в его восточной части или у кромки паковых льдов в Чукотском море достигают в июле. Большинство из них совершает эти переходы по разводьям между припаем и дрейфующими льдами. Такова по крайней мере гипотеза. Сто лет назад, когда охота на гренландских китов достигала своего пика, летом больше всего животных били именно вдоль кромки летнего льда. Доподлинно неизвестно, чем питаются гренландские киты, но, вероятно, главную их пищу составляют рачки копеподы. Они намного мельче криля, но гренландский кит отфильтровывает их из воды с помощью чрезвычайно тонких пластин. Возникает вопрос: каким образом популяция гренландского кита сохраняется в местообитании, известном своей малой продуктивностью? А что продуктивность летней житницы китов - моря Бофорта - скудна, доказывается тем, что в его водах тюлени и киты широко представлены только одним видом, первые - маленькой кольчатой нерпой (Phoca hispida), вторые - белухой. Может быть, киты питаются более эффективным способом, чем большинство морских млекопитающих. Или мы ошибочно переносим на гренландских китов сведения, относящиеся к другим гладким китам. Большинство из них кормятся в основном летом, а в остальные времена года едят мало или вообще постятся. Возможно, эти киты питаются круглый год, используя неизвестные нам пищевые ресурсы. Не исключено, например, что они находят себе пропитание в Беринговом море и зимой, когда другие крупные киты, по нашим предположениям, едят мало. Гренландские киты и для спаривания выбирают странное время. Скорее всего, они спариваются летом, а не поздней осенью и зимой, как другие гладкие киты.

Итак, гренландские киты - самые достопримечательные из всех гладких китов, они как бы воплощают в себе исключения из «правил», коим подчинена жизнь их собратьев. Они не мигрируют в теплые воды, как, например, полосатики: синие киты (Balaenoptera musculus), достигающие в длину свыше 30 м, и двадцатиметровые финвалы (Balaenoptera physalus). Полосатики в полярных и субполярных морях лишь кормятся, щениться же и выкармливать детенышей отправляются в моря умеренных и даже тропических широт. Другой вид китов, серый кит (Eschrichtius gibbosus), также находится в Арктике только летом. Зимует он в тропических бухтах Мексики, там выводит и выкармливает молоком потомство. Южный кит (Eubalaena glacialis) (Южный кит имеет 3 подвида (иногда они рассматриваются как отдельные виды): японский кит (Eubalena glacialis sieboldi), бискайский кит (Е. g. glacialis) и австралийский кит (Е. g. australis). Ареалы их широко разделены тропическим поясом и материками. У берегов Южной Африки встречается австралийский кит, он распространен в южном полушарии от 15 до 60° ю. ш., главным образом между 30 и 50° ю. ш. Биология всех трех подвидов очень сходна. Сейчас промысел южных китов запрещен международным соглашением. - Прим. ред. ) , ближайший родственник гренландского кита, фактически представляет собой прибрежный вид и обитает в умеренных зонах, в субполярных же морях он придерживается окраин, а сейчас встречается вообще чрезвычайно редко, за исключением Южной Африки, где его популяция сильно возросла.


Группа белух в холодных водах Канады.Белухи часто собираются в большие группы, насчитывающие подчас сотни животных. Это, однако, случайные объединения, лишенные четкой социальной организации. Белухи, можно сказать, обладают "хорошими вокальными данными", за что заслужили прозвище морских канареек

Нарвал и белуха состоят в близком родстве, оба живут в водах с преобладанием льда. Они принадлежат к зубатым китам, хотя взрослый нарвал не имеет зубов. Вместо них с левой стороны у него развивается могучий, спирально закрученный бивень, торчащий вперед на 2-3 м. Обычно нарвал имеет один бивень, но встречаются и экземпляры с двумя бивнями. И тут мы сталкиваемся с очень странным нарушением одного биологического закона. Он гласит, что всем позвоночным присуща двусторонняя симметрия, т. е. одна сторона тела является зеркальным отражением другой. Но если у нарвала вырастают два бивня, они все равно оба помещаются слева (Правый бивень у самца и оба бивня у самки скрыты в челюстях. Лишь крайне редко они развиваются у самцов и у самок. Нарвалы приспособились жить в разводьях среди льдов, и их не страшит опасность задохнуться, когда замерзают полыньи: самцы разламывают лед, нанося бивнем фронтальные удары снизу. Через пробитое отверстие дышат все члены стада. В случае поломки бивня его зубной канал закрывается костной пробкой. - Прим. ред. ) . Что касается функции бивней, то недавние исследования подтвердили прежние гипотезы. Бивнем обладают исключительно самцы, используют они его главным образом при демонстрационном поведении для привлечения самок и только очень редко - в драках. Такую же роль играют рога у оленей. Различные наблюдатели не раз видели, как нарвалы демонстрируют свои бивни и даже скрещивают их над водой, подобно тому как гладиаторы скрещивали мечи. А может быть, как и у моржей, подобная демонстрация является способом установления нарвалами социальной иерархии. Белухи, или белые киты, известны намного лучше. Они не проявляют такой беззаветной преданности льду, как гренландские киты или нарвалы. Белух можно встретить значительно дальше на юг, например, близ устья канадской реки Св. Лаврентия или у берегов Южной Аляски. Детеныши при рождении имеют серовато-синий оттенок, ближе к зрелости они белеют. Основная пища белух - рыба, но они поглощают и различных беспозвоночных. Известно, что, используя приливные течения, белухи поднимаются вверх по рекам вслед за лососевыми и уничтожают этих ценных промысловых рыб. В далеком прошлом, когда и белух, и лососей было больше, чем рыболовов, это никому не мешало, теперь же гастрономические наклонности белухи вызывают возмущение. Но так как мало кому хочется убивать красивых доверчивых белух, было найдено компромиссное решение: во избежание столкновений между китами и рыбаками проигрывать в местах скоплений белух запись звуков, издаваемых под водой косатками (Orcinus огса). Киты в ужасе обращаются в бегство, а лососи достаются рыбакам.


Косатки, или киты-убийцы, также живут в полярных водах. Это самые большие из дельфинов: наиболее крупные самцы вырастают до 8 м, их вес - 5 - 6 т. Косатки встречаются в морях умеренного пояса, но, вероятно, наибольшее поголовье вида сосредоточено в Южном океане и в северной части Тихого, до самого Берингова моря (Косатки - настоящие космополиты: живут во всех океанах, от Арктики до Антарктиды, где далеко заходят в плавучие льды. В СССР наблюдаются в таких арктических морях, как Карское и Восточно-Сибирское (Чаунская губа). - Прим. ред. ) . Косатку очень украшают белые пятна, расположенные над глазом, ниже спинного плавника и на брюхе. Убийцей ее прозвали недаром: она способна убивать самых крупных китов, тюленей, дельфинов, сивучей, хотя питается преимущественно рыбой. Косатки большими стаями, вроде волчьих, нападают на гладких китов, выедают у своих жертв язык, сдирают с них жир, вырывают из тела куски мяса. Мелких тюленей они заглатывают целиком. В то же время в неволе косатки проявляют недюжинный интеллект и редкую доброту.
«Наземные» морские млекопитающие

Природа не скупится на отступления от общих правил, в этом убеждает пример белого медведя. Он в два раза тяжелее тигра, а по величине уж во всяком случае не уступит бурому медведю с Аляски. В отличие от других животных он почти не боится людей, даже иногда на них нападает. Впрочем, чаще всего он нас просто не замечает. Белые медведи появились на Земле во время ледниковой эпохи, их предками были бурые медведи типа гризли. Морским это млекопитающее считается только потому, что больше всего любит рыскать по морскому льду, выискивая добычу - тюленей и слабых моржат (В противоположность другим видам белых медведей, ведущим наземный образ жизни, белый медведь (Ursus maritimus) является типичным обитателем Арктики, лишь кое-где заходящим в материковые тундры. Необычайно густая плотная шерсть отлично защищает тело от холода и намокания в ледяной морской воде, подошвы его ног сплошь покрыты шерстью, уши едва выдаются над поверхностью волосяного покрова. Важную приспособительную роль играет мощный слой подкожного жира. - Прим. ред. ). В его анатомии и физиологии не произошло сколько-нибудь важных изменений для приспособления к жизни в воде. И тем не менее, пища белого медведя состоит в основном из морских млекопитающих. Иногда он берет и рыбу, но чаще всего его жертвой становятся те морские животные, которые живут на морском льду и не успевают достаточно быстро скрыться при появлении медведя. Он отыскивает норы кольчатой нерпы среди нагромождений обломков припая, вытаскивает оттуда детенышей и пожирает. В другое время года он подолгу просиживает у продушины в ожидании взрослых кольчатых нерп. Стоит нерпе всплыть на поверхность, чтобы набрать воздуха, как он одним ударом своей могучей передней лапы, вооруженной когтями, выбивает ее на лед. Кольчатая нерпа считается основной пищей белого медведя, но это справедливо лишь отчасти, так как наблюдения за медведями велись чаще всего около берегов, где нерпы преобладают и где их легче всего увидеть.

Белые медведи встречаются во множестве и вдали от берегов, иногда они заходят за сотни миль от суши. Кольчатые нерпы редко отдаляются на такие расстояния от припайного льда, и, естественно, возникает вопрос: чем же тогда питаются медведи? Моржи - самые крупные из всех ластоногих Арктики. Обитают они среди морских льдов, держатся большими стадами, с чего бы, казалось, им бояться белого медведя, который вряд ли осмелится напасть на животных огромных размеров, вооруженных, к тому же, могучими бивнями? Оказывается, что на льду морж становится трусливым, в отличие от тюленей Антарктики, где отсутствуют крупные наземные хищники. При виде приближающегося человека или подкрадывающегося медведя им овладевает паника, он в ужасе кидается в воду, часто забывая про своего детеныша. Белый медведь нападает на группу моржей с явным расчетом запугать их и, если ему повезет, хватает замешкавшегося моржонка за задние ласты. Убив его, хищник делает надрез на затылке и выедает жир и мясо, постепенно выворачивая наизнанку шкуру, которая остается единственной, кроме костей, уликой на месте преступления.

Наши братья морские
Тюлени и моржи принадлежат к млекопитающим, что не мешает им сохранять тепло в полярных морях, опускаться под воду на тысячи метров - по крайней мере на один-два километра, -находиться на этой глубине более часа. И все это потому, что они способны переносить высокую кислотность крови, вызываемую накоплением побочных продуктов метаболизма, и запасать кислород в больших количествах, чем можем мы. Их терморегуляция, иными словами контроль над поступлением и отдачей тепла, представляет собой сложный механизм, который предусматривает определенные размеры и форму тела, изоляцию, способы теплообмена, сосудистый контроль над потоком крови, поведение животного. Если этот механизм действует исправно, моря, даже полярные, кажутся им теплыми, но любые неполадки в нем вызывают у животных болезненную реакцию, вплоть до смертельного исхода. Если, к примеру, пищи мало и тело животного недостаточно защищено слоем жира, оно не может ни сопротивляться холоду, ни вырабатывать тепло в необходимом для существования количестве. Тех детенышей, что появляются на свет к концу сезона выведения потомства, приходится преждевременно отнимать от груди, и они, не успев набрать нужного веса, не доживают до лета.
Непонятным образом, просто чудом, может быть именно благодаря сходству с нами, морские млекопитающие умудряются жить - даже процветать! - в самой враждебной на Земле среде - в полярных морях.